|
|
|
|
|
GENETIK |
Molekulargenetik: Das Jacob-Monod-Modell |
|
|
|
Nach den Forschungen von Francois Jacob und Jacques Monod und dem von den beiden Forschern aufgestellten Schema vollzieht sich die Regulation der Proteinsynthese nach einem bestimmten Mechanismus. Das Jacob-Monod-Modell war der erste entdeckte und entschlüsselte Genregulationsmecahnismus. Inzwischen sind zahlreiche andere Modelle dazu gekommen.
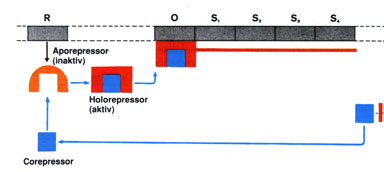
Abb.: Das Regulatorgen R produziert ein Molekül, einen inaktiven
Aporepressor. Wenn sich dieser Aporepressor mit einem Corepressor
(einem Substrat) verbindet, so entsteht ein aktiver Holorepressor,
der den Operator verschließt. Die Ablesung der Strukturgene S1,
S2, ... usw. wird dadurch verhindert. (Der rote Strich symbolisiert
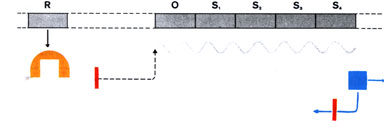
die Ableseblockade). Es muss neben den Genen für die Synthese der Enzyme noch besondere Gene geben, die für die Regulierung der Aktivität dieser Enzymgene verantwortlich sind. Durch Genkartierung beim Bakterium Escherichia coli wurde festgestellt, dass die Gene, die die Struktur eines spezifischen Proteins bestimmen, stets unmittelbar nebeneinander liegen; sie heißen daher Strukturgene, werden auch stets gemeinsam "abgelesen", und bei der Transkription entsteht eine einzige m-RNA. Die Ablesung der Strukturgene wird von dem Protein eines weiteren Gens, dem Regulatorgen, gesteuert. Dieses Protein hemmt die Ablesung der Strukturgene und wird als Repressor bezeichnet. Der Repressor wirkt spezifisch auf die ihm jeweils zugeordnete Strukturgene, tritt also mit ihnen in Wechselwirkung. Dies wird durch ein benachbartes DNA-Stück, das als Operator bezeichnet wird, erreicht. Neben diesem liegt ein weiterer kurzer DNA-Abschnitt, der als Anlagerungsstelle
für die RNA-Polymerase dient. Operator und Strukturgene zusammen
bilden ein Operon. Wenn sich ein Repressormolekül an den Operator
anlagert, kann die RNA-Polymerase nicht gebunden werden. Damit wird die
Synthese der m-RNA an den Strukturgenen unterbunden. (Zur Erinnerung:
Die m-RNA transportiert den genetischen Code aus dem Zellkern ins Plasma,
wo die Proteine synthetisiert werden).
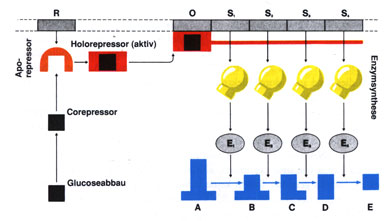
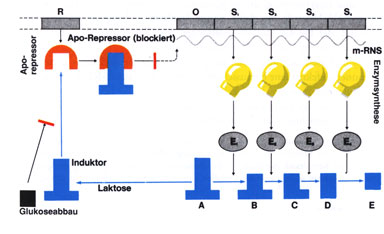
Vor allem bei der Synthese von Enzymen für abbauende Stoffwechselreaktionen erfolgt die Steuerung der Genaktivität durch Substratinduktion; d. h. die Auslösung der Enzymsynthese wird durch das Substrat bewirkt, das als Induktor fungiert. Eine andere Möglichkeit der Regulation ist die Hemmung vom Endprodukt her: Wird das Endprodukt in der Zelle nicht weiterverwertet und reichert sich daher an, wirkt es als Corepressor. Dadurch wird der Repressor in seiner Form verändert, lagert sich an den Operator und verhindert so die weitere Transkription der Strukturgene. |
|
|
Vorarlberger Bildungsserver |