
|
|
|
|
|
BOTANIK |
Der Ablauf der Photosynthese |
|
 |
Die lichtabhängigen Reaktionen Die Untersuchungen über den Einfluss der Photosyntheseleistung haben gezeigt, dass es bei der Photosynthese Licht- und Dunkelreaktionen gibt. Vertiefte Erkenntnisse dieser beiden Reaktionsabläufe erhält man, wenn man die an der Reaktion beteiligten Strukturen aus der Pflanze isoliert und weiter untersucht. So zeigte sich, dass der ATP-Gehalt in den Chloroplasten bei Belichtung ansteigt und dass dabei bestimmte Substanzen reduziert und oxidiert werden. An Thylakoiden, die aus isolierten, osmotisch Da diese Reaktion auch bei Abwesenheit von Kohlenstoffdioxid ablief, war gleichzeitig nachgewiesen, dass der freigesetzte Sauerstoff aus dem Wasser stammt. Der Sauerstoff entsteht durch Spaltung des Wassers unter Lichteinfluss. Man spricht von der Photolyse des Wassers, die nach ihrem Entdecker auch Hill-Reaktion genannt wird.
Als natürlicher Elektronenakzeptor tritt in der Zelle das NADP+
(Nicotinamid-Adenin-Dinukleotid-Phosphat) auf. Abb.: Reduktion von NADP Das im Reaktionszentrum aktive Chlorophyll a löst die photochemische Reaktion aus. Diese Reaktion ist im Wesentlichen ein Elektronentransport. Bei diesem Transport geht ein angeregtes Elektron auf das Akzeptormolekül NADP+ über. Das Chlorophyll a wird dabei oxidiert, der Akzeptor reduziert, indem er Wasserstoffionen aus der Photolyse aufnimmt. Im Molekül hinterlassen diese Elektronen ein "Loch". Dieses Loch kann von anderen freien Elektronen wieder aufgefüllt werden. Der Gesamtprozess der Lichtreaktion lässt sich in drei Teilreaktionen
gliedern: Die einzelnen Teilschritte der Lichtreaktion lassen sich im so genannten "Zick-Zack-Schema" (Abb. unten) darstellen. In diesem Schema sind die am Elektronentransport beteiligten Überträgersubstanzen ihrem Energieniveau entsprechend eingeordnet.
Durch Licht angeregte Chlorophyllmoleküle des Photosystems II entziehen über eine manganhaltige Überträgersubstanz Z dem Wasser Elektronen. Das Wasser wird zu Sauerstoff oxidiert, dabei werden ebenfalls H-Ionen frei (Photolyse). Die Elektronen werden über eine Kette von Überträgersubstanzen (Substanz Q, Plastochinon, Cytochrom b, Cytochrom f, Plastocynanin) "bergab" an das Photosystem I transportiert, das dadurch vom angeregten Zustand in den Grundzustand zurückkehrt. Bei erneuter Anregung durch Licht werden die Elektronen vom System I über das Ferredoxin und das Flavoprotein geleitet und zur Reduktion von NADP+ benutzt. Man spricht von einem offenkettigen oder nichtzyklischen Transport. Der Übergang der Photosysteme vom Grundzustand in den angeregten Zustand ist eine endergonische Rektion. Beim Bergabfließen der Elektronen handelt es sich jeweils um exergonische Reaktionen, die Energie freisetzen. Die Photolyse des Wassers und die Reduktion von NADP+ binden einen Teil der absorbierten Lichtenergie. Ein anderer Teil dieser Energie wird zur Bildung von ATP genutzt. Die Synthese von ATP bezeichnet man als Phosphorylierung im Licht oder als nichtzyklische Photophosphorylierung. Neben dem offenkettigen, nichtzyklischen Elektronentransport existiert noch ein zyklischer Transport. Dieser findet zum Beispiel dann statt, wenn man nur das Licht der Wellenlänge 700 nm verwendet. An diesem Transport kann also nur das System I beteiligt sein. Dabei werden vom angeregten P700 Elektronen über das Ferredoxin wieder zum P700 zurückgeleitet. Dieser Vorgang ist im Gegensatz zum nichtzyklischen Transport nicht mit der Bildung von Sauerstoff und NADPH+ H+ gekoppelt. Allerdings entsteht bei diesem zyklischen Transport, ebenso wie beim nichtzyklischen Transport, ATP. Man bezeichnet diesen Vorgang als zyklische Photophosphorylierung. Die bei der Lichtreaktion geleistete Arbeit besteht darin, dass Elektronen des Wassers letztlich auf NADP+ übertragen und damit auf ein hohes Energieniveau gehoben werden. Dieses Anheben ist ein endergoner Prozess, bei dem eine Redoxpotentialdifferenz von 1,13 Volt überwunden, d. h. ein Energiebetrag von G = 218 kJ pro Mol NADPH + H+ aufgebracht werden muss. Diese Energie wird durch vier Lichtquanten geliefert. Sie treiben zwei in Serie geschaltete "Elektronenlifts" - nämlich die beiden Photosysteme - an. Das Zusammenwirken der beiden Systeme macht es möglich, dass die Elektronen unterwegs ein Stück "bergab" laufen und dabei ATP gebildet wird. Beim Elektronentransport und bei der ATP-Synthese handelt es sich um Reaktionen, die in den Thylakoidmembranen der Chloroplaste in genau abgegrenzten Bereichen auftreten. 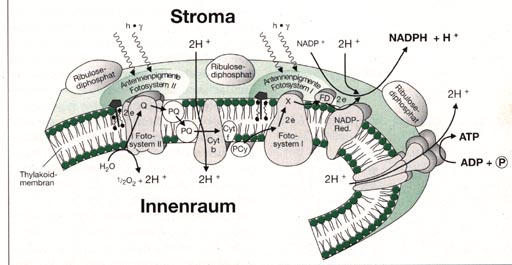
Abb.: Modell zur räumlichen Anordnung der Redoxsysteme in der Thylakoidmembran Die lichtunabhängigen Reaktionen Der Eintritt des Kohlenstoffdioxid erfolgt bei den Landpflanzen zum größten Teil über die Stomata. Innerhalb des Blattes diffundiert das atmosphärische Kohlenstoffdioxid in das interzelluläre Durchlüftungsgewebe und löst sich in dem Wasser, das sich im Bereich der Zellwände befindet. Das bei der Zellatmung entstehende Kohlenstoffdioxid kommt hinzu. Kohlenstoffdioxid liegt im Wasser teils gelöst, teils in Form von Kohlensäure, Carbonat- oder Bikarbonat-Ionen vor. Aus dem Zellwandbereich gelangt das gelöste Kohlenstoffdioxid in
die nur wenige Mikrometer entfernten Chloroplasten. Die Diffusion wird
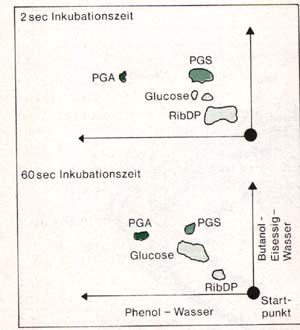
dabei durch die Plasmaströmung der Zellen unterstützt. Abb.: Autoradiogramm einiger Photosyntheseprodukte nach 2 Sekunden und nach 60 Sekunden Belichtungszeit bei Chlorella Die Aufklärung des weiteren Weges des Kohlenstoffdioxids und die Bildung von Glucose ist in erster Linie dem Einsatz von radioaktiv markierten Isotopen zu verdanken. Die entscheidenden Versuche wurden von CALVIN und seinen Mitarbeitern an einzelligen Algen durchgeführt. Bei solchen Versuchen gibt man der Algensuspension radioaktiv markiertes Kohlenstoffdioxid zu (14C02 oder NaH14C03). Einige Sekunden oder Minuten nach der Eingabe des 14CO2 wird der Ansatz jeweils durch Eingießen in siedenden Alkohol abgetötet. Die Dauer der Kohlenstoffdioxid-Fixierungsperiode wird bei den einzelnen Versuchen variiert. Die löslichen Inhaltsstoffe werden extrahiert und chromatographisch getrennt. Die nachzuweisenden Mengen sind jedoch für chemische Nachweisreaktionen zu klein. Aus diesem Grunde bringt man das Chromatogramm im Dunkeln in engen Kontakt mit einem unbelichteten Röntgenfilm. Die hochempfindliche Schicht des Filmes wird durch die radioaktive Strahlung des 14C geschwärzt. Die so ermittelten Substanzen werden anschließend identifiziert. Man bezeichnet dieses Verfahren als Autoradiographie. Nach 2 Sekunden Fixierungszeit (siehe Abb. oben) ist der größte Teil des radioaktiv markierten Kohlenstoffs in Form von Phosphoglycerinsäure festgelegt. Es ist das erste stabile Zwischenprodukt, das man inzwischen bereits nach 0,4 s nachweisen kann. Alle übrigen Produkte entstehen durch biochemische Umwandlung der Phosphoglycerinsäure. Man nimmt an, dass sie durch den Zerfall eines instabilen Moleküls mit 6 C-Atomen (C6-Körper) gebildet wird. Diese Verbindung entsteht durch Anlagerung des Kohlenstoffdioxid an einen C5 -Körper, das Ribulosediphosphat. Durch das Ribulosediphosphat wird das Kohlenstoffdioxid in die Dunkelreaktion eingeschleust. Nach rund 60 s hat sich ein Gleichgewicht zwischen Phosphoglycerinsäure und Ribulosediphosphat eingestellt. Dies ist ein Anzeichen dafür, dass es sich bei dem Gesamtvorgang der Kohlenstoffdioxid-Verarbeitung um einen Kreisprozess handelt, den man als Calvin-Zyklus bezeichnet.
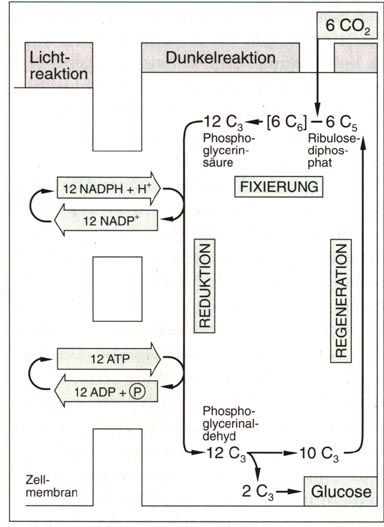
Abb. links: Fixierung und Reduktion des Kohlendioxids sowie Regeneration des Ribulose-diphosphats im Calvin-Zyklus Die ständige Regeneration des Ribulosediphosphats geschieht in einer komplizierten Folge von chemischen Reaktionen. Hierzu dient der größte Teil des Phosphoglycerinaldehyds. Von 12 Molekülen werden zwei zu Glucose umgebaut, die restlichen 10 C3-Körper dienen zum Wiederaufbau von 6 C5-Körpern, dem Ribulosediphosphat, das dann wiederum als Kohlenstoffdioxid-Akzeptor auftreten kann. Der Calvin-Zyklus erfüllt zwei Aufgaben. ibulosediphosphat. Diese
beiden Teilprozesse finden in den Stromathylakoiden der Chloroplasten
statt. Die Aufnahme des körperfremden Kohlenstoffdioxids und seine
Umwandlung in körpereigenes Substrat werden auch als Kohlenstoff-Assimilation
bezeichnet.
In den lichtabhängigen Reaktionen müssen demnach 12 Wassermoleküle
durch Photolyse zerlegt werden und 24 Setzt man den Energieaufwand alle Photosynthetischen Reaktionen in Relation zu der in der Glucose gebundenen chemischen Energie, so ergibt sich ein Wirkungsgrad von rund 30 %. Spezialisten im C02-Gaswechsel: Die C4 Pflanzen Hält man eine Maispflanze und eine Sojabohnenpflanze in einem gasdichten Behälter zusammen, so wächst die Maispflanze heran, während die Sojabohnenpflanze schon nach wenigen Tagen abstirbt. Ursache dafür ist, dass die Maispflanze im Licht viel wirkungsvoller Kohlenstoffdioxid aufnehmen und fixieren kann als die Sojabohnenpflanze und ihr im "Wettbewerb" um das für die Photosynthese so wichtige Ausgangsprodukt CO2überlegen ist.
Der Mais zeigt schon rein anatomisch eine charakteristische zelluläre Anordnung seiner Blattgewebe. Im Gegensatz zu den meisten Landpflanzen besitzt diese Pflanze Leitbündel, die von einer chloroplastreichen Leitbündelscheide kranzförmig umgeben sind. Die Chloroplasten der Leitbündelscheidezellen enthalten große Stärkekörner, viel Stromathylakoide und nur wenig Granathylakoide. Die Leitbündelscheidezellen sind wiederum umgeben von Mesophyllzellen, deren Chloroplasten den üblichen Aufbau zeigen. Man spricht von einem Chloroplastendimorphismus. Diese speziellen anatomischen Verhältnisse gehen einher mit besonderen physiologischen Gegebenheiten. In den Chloroplasten der Mesophyllzellen wird das bei der Zellatmung produzierte ebenso wie das aus der Luft aufgenommene Kohlenstoffdioxid durch Phosphoenolbrenztraubensäure, einen C3-Körper, fixiert und über Oxalessigsäure in einen C-Körper umgewandelt, das Malat (Apfelsäure). Entsprechend werden diese Pflanzen C4 Pflanzen genannt. Das Malat wird durch feine Plasmakanäle aus den Mesophyllzellen in die Leitbündelscheidezellen transportiert und dort decarboxiliert. Das dabei gebildete Kohlenstoffdioxid steht der Pflanze dort in großer Menge für die Reduktion im Calvin-Zyklus zur Verfügung, der ausschließlich in den Leitbündelscheidezellen abläuft. Fixierung des Kohlenstoffdioxids und seine Reduktion im Calvin-Zyklus erfolgen räumlich getrennt. 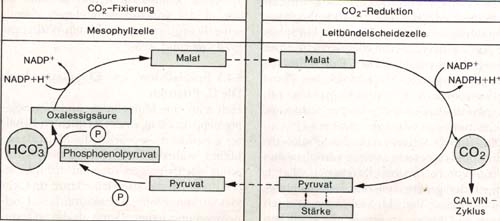
Abb.: Räumliche Trennung der CO2-Fixierung und der CO2-Reduktion bei C4-Pflanzen Bei Bildung des Malat wird das durch die Spaltöffnungen eindiffundierende Kohlenstoffdioxid aus dem Konzentrationsgefälle "Außenluft-Blattinneres" herausgenommen. Auf diese Weise können im Licht bei geöffneten Spalten große Mengen Kohlenstoffdioxid in das Blatt eindringen. Man kann den C4-Mechanismus daher auch als eine lichtgetriebene "C02 Pumpe" zur Konzentrierung des Kohlenstoffdioxids für den Calvin-Zyklus auffassen. Durch die Zwischenspeicherung im Malat ist das Kohlenstoffdioxid in der Pflanze so stark konzentriert, dass es bei Belichtung in der Regel nicht als begrenzender Faktor auftreten kann. Seine höhere Verfügbarkeit macht die C4 Pflanzen zu Photosynthetischen Hochleistungspflanzen. Neben dem Mais zählt auch das Zuckerrohr zu dieser Gruppe. Die Fixierung von Kohlenstoffdioxid auf dem C4 Wege erfolgt also viel intensiver als bei einer Pflanze, bei der das von außen aufgenommene Kohlenstoffdioxid direkt in Form von Phosphoglycerinsäure - einem C3 Körper - im Calvin-Zyklus fixiert wird. Man bezeichnet letztere als C3 Pflanze. Während durchschnittliche C3 Pflanzen etwa 800 g Wasser transpirieren, um 1 g Trockenmasse zu bilden, benötigen C4-Pflanzen dafür meist weniger als die Hälfte. Der C4 Weg ist auch eine Möglichkeit Wasser zu sparen. Es ist daher nicht verwunderlich, dass die Geröllhalden im "Tal des Todes" in Kalifornien, einem Gebiet mit einer Durchschnittstemperatur im Juli von 39 °C nur von Pflanzen besiedelt werden, die den C4 Mechanismus besitzen. Über einen grundsätzlich ähnlichen Mechanismus der C02 Fixierung verfügen viele Pflanzen, deren Standortbedingungen gekennzeichnet sind durch starke Sonneneinstrahlung, hohe Temperaturen am Tage und extremen Wassermangel. Pflanzen, die unter solchen Bedingungen leben, müssten eigentlich am Tage ihre Stomata weit öffnen, so dass Kohlenstoffdioxid ins Blattinnere diffundieren kann. Dies führt jedoch zu einem lebensbedrohenden Wasserverlust. Der C4 Mechanismus stellt für diese Pflanzen eine Möglichkeit dar, dieses "Dilemma" zu mildern. Bei den Blättern dieser Pflanzen, zu denen vor allem die Sukkulenten (z. B. Fetthenne und Sempervivum-Arten) gehören, beobachtet man während der Nacht eine starke Anhäufung von organischen Säuren, vor allem von Malat. Diese Pflanzen öffnen ihre Stomata während der Nacht, nehmen viel Kohlenstoffdioxid auf und speichern es im Malat. Die Speicherung erfolgt in den Vakuolen der Blattzellen. Dies trägt dazu bei, dass die Blätter sich fleischig anfühlen. Die deponierte Säure wird am Tage wieder abgebaut. Die Säuremenge in den Blättern ist entsprechend starken tageszeitlichen Schwankungen unterworfen. Man spricht von einem diurnalen Säurerhythmus. Durch die intensive Decarboxlierung des Malat am Tage kommt es zu einer hohen C02-Konzentration im Blattinnern. Dies verursacht zum einen den Verschluss der Spalten und verhindert zum anderen den Verlust des lebenswichtigen Wassers. Fixierung und Reduktion des Kohlenstoffdioxids sind bei diesen Pflanzen zeitlich getrennt. Bilder: Kroll, Demmer, Thies: "Stoffwechsel", Serie Biologie, Westermann Wien |
|
|
Welt der Biologie |
| Anatomie/Physiologie | Botanik | Cytologie | |
| Evolution | Genetik | Humanbiologie | Ökologie |
| Sexualbiologie | Zoologie | Geschichte | Texte, Referate |
| Sehenswert | Kontakt | Physik |
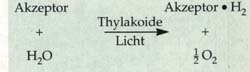 aufgebrochenen
Chloroplasten stammten, konnte nachgewiesen werden, dass diese bei Belichtung
Photochemisch aktiv sind. Weiterhin gelang es bei diesen Versuchen künstliche
Elektronenakzeptoren, etwa den Farbstoff Dichlorphenolindophenol (DCPIP),
zu reduzieren und dabei Sauerstoff freizusetzen.
aufgebrochenen
Chloroplasten stammten, konnte nachgewiesen werden, dass diese bei Belichtung
Photochemisch aktiv sind. Weiterhin gelang es bei diesen Versuchen künstliche
Elektronenakzeptoren, etwa den Farbstoff Dichlorphenolindophenol (DCPIP),
zu reduzieren und dabei Sauerstoff freizusetzen.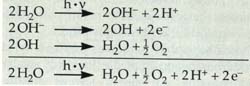 Durch
die Elektronenabgabe wird das OH-Ion zum Radikal. Zwei OH-Radikale reagieren
miteinander zu Wasser und Sauerstoff. Der entstehende Sauerstoff wird
im weiteren Verlauf der Photosynthese nicht mehr gebraucht. Wichtig sind
jedoch die gebildeten Wasserstoffionen und die dem Wasser entzogenen Elektronen.
Durch
die Elektronenabgabe wird das OH-Ion zum Radikal. Zwei OH-Radikale reagieren
miteinander zu Wasser und Sauerstoff. Der entstehende Sauerstoff wird
im weiteren Verlauf der Photosynthese nicht mehr gebraucht. Wichtig sind
jedoch die gebildeten Wasserstoffionen und die dem Wasser entzogenen Elektronen.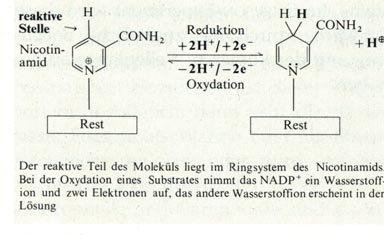 Belichtet
man nämlich isolierte Chloroplasten bei Abwesenheit von CO2
und Anwesenheit von ADP, anorganischem Phosphat und NDAP+,
so entstehen ATP, NADPH+, H+ und Sauerstoff. Aufgrund
solcher und ähnlicher Befunde lässt sich ein vereinfachtes Modell
der Lichtreaktion entwickeln.
Belichtet
man nämlich isolierte Chloroplasten bei Abwesenheit von CO2
und Anwesenheit von ADP, anorganischem Phosphat und NDAP+,
so entstehen ATP, NADPH+, H+ und Sauerstoff. Aufgrund
solcher und ähnlicher Befunde lässt sich ein vereinfachtes Modell
der Lichtreaktion entwickeln.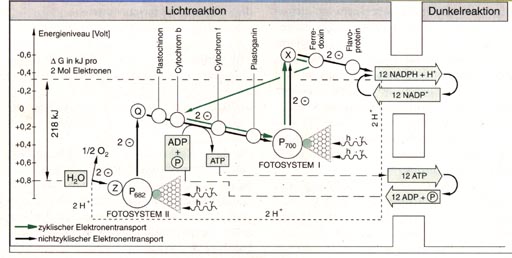
 Durch
Änderung der Fixierungszeiten hat man die wichtigsten Syntheseschritte
dieses Kreisprozesses aufklären können. Zunächst wird die
Phosphoglycerinsäure durch das in den Lichtreaktionen gebildete ATP
aktiviert und durch das ebenfalls aus dieser Reaktion stammende NADPH+
H+ zu Phosphoglycerinaldehyd reduziert. Mit dem Phosphoglycerinaldehyd
ist die energiereiche Stufe der Kohlenhydrate erreicht. Durch Zusammenlagerung
von zwei Molekülen Phosphoglycerinaldehyd entsteht Fructose-1,6-Diphosphat.
Dieser C6 Körper wird nun schrittweise zu Glucose umgebaut.
Durch
Änderung der Fixierungszeiten hat man die wichtigsten Syntheseschritte
dieses Kreisprozesses aufklären können. Zunächst wird die
Phosphoglycerinsäure durch das in den Lichtreaktionen gebildete ATP
aktiviert und durch das ebenfalls aus dieser Reaktion stammende NADPH+
H+ zu Phosphoglycerinaldehyd reduziert. Mit dem Phosphoglycerinaldehyd
ist die energiereiche Stufe der Kohlenhydrate erreicht. Durch Zusammenlagerung
von zwei Molekülen Phosphoglycerinaldehyd entsteht Fructose-1,6-Diphosphat.
Dieser C6 Körper wird nun schrittweise zu Glucose umgebaut.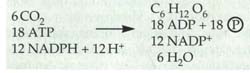 Für
die Herstellung eines Glucose-Moleküls in der lichtunabhängigen
Dunkelreaktion werden benötigt:
Für
die Herstellung eines Glucose-Moleküls in der lichtunabhängigen
Dunkelreaktion werden benötigt: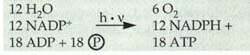 Elektronen
über die Photosysteme fließen, was zusammen 48 Lichtanregungsprozesse
erforderlich macht.
Elektronen
über die Photosysteme fließen, was zusammen 48 Lichtanregungsprozesse
erforderlich macht.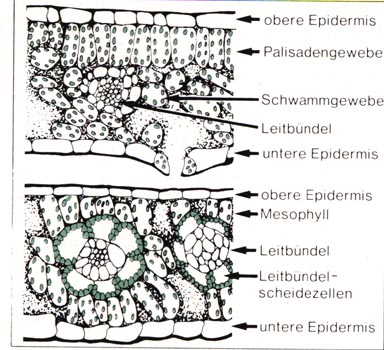 Abb.:
Blattquerschnitt einer C3-Pflanze (Helleborus) und einer C4-Pflanze
(Mais)
Abb.:
Blattquerschnitt einer C3-Pflanze (Helleborus) und einer C4-Pflanze
(Mais)